
Artigo original
FUNGAL POLYSACCHARIDES AND THEIR HYPOGLYCEMIC POTENTIAL TO ACT AS AN ADJUVANT IN THE TREATMENT OF DIABETES MELLITUS
Polissacarídeos fúngicos e seu potencial hipoglicemiante para atuar como adjuvante no tratamento do diabetes mellitus
https://doi.org/10.18593/evid.32561
Recebido em 19 de fevereiro de 2023 | Aceito em 12 de abril de 2023
Gabriel Macêdo Figueroa* Danilo Nascimento Costa† Alana Caise dos Anjos Miranda‡ Gildomar Lima Valasques Júnior§ Tátilla Putumujú Santana Mendes¶ Pâmala Évelin Pires Cedro**
Danilo Nascimento Costa† Alana Caise dos Anjos Miranda‡ Gildomar Lima Valasques Júnior§ Tátilla Putumujú Santana Mendes¶ Pâmala Évelin Pires Cedro**
Abstract:
Diabetes mellitus is considered a serious public health problem because it affects millions of individuals and causes numerous complications. The constant adverse effects of conventional therapies for diabetes mellitus have stimulated research that seeks new forms of treatment, prevention and control. Polysaccharides obtained from fungi are promising as adjuvants for the control of diabetes mellitus. This study is a literature review that sought to evaluate the hypoglycemic properties of polysaccharides obtained from fungi and understand the main mechanisms of action involved. The main mechanisms reported in the studies resulting in a decrease in glycemia were the inhibition of digestive enzymes such as α-amylase and α-glucosidase, action on the IRS1, PI3K, JNK1 signaling pathways, antioxidant, anti-inflammatory, hypolipidemic capacity and stimulation of the intestinal microbiota.
Keywords: Biotechnology. Fungi. Hypoglycemics.
Resumo:
Diabetes mellitus é considerado um grave problema de saúde pública, pois afeta milhões de indivíduos e causa inúmeras complicações. Os constantes efeitos adversos das terapias convencionais para diabetes mellitus têm estimulado pesquisas que buscam novas formas de tratamento, prevenção e controle. Polissacarídeos obtidos de fungos são promissores como adjuvantes no controle do diabetes mellitus. Este estudo é uma revisão de literatura que buscou avaliar as propriedades hipoglicemiantes de polissacarídeos obtidos de fungos e compreender os principais mecanismos de ação envolvidos. Os principais mecanismos relatados nos estudos que resultaram na diminuição da glicemia foram a inibição de enzimas digestivas, como α-amilase e α-glicosidase, ação nas vias de sinalização IRS1, PI3K, JNK1, antioxidante, anti-inflamatória, capacidade hipolipidêmica e estimulação da microbiota intestinal.
Palavras-chave: Biotecnologia. Fungos. Hipoglicemiantes.
@Autor correspondente: Mestre em Bioquímica e Biologia Molecular, Universidade Estadual do Sudoeste da Bahia, Avenida José Moreira Sobrinho, Jequiézinho, Jequié, BA, Brasil; https://orcid.org/0000-0002-2888-1140; pamalaevelinpires@hotmail.com.
1 INTRODUCTION
Diabetes mellitus is a chronic metabolic disease characterized by a persistent increase in blood glucose1, which occurs due to a deficiency in the production or action of insulin, configuring type 1 diabetes and type 2 diabetes, respectively. Diabetes is a major health problem that reaches alarming levels year after year2.
About 463 million individuals live with diabetes worldwide, and it is estimated that in 2030 and 2045 the number of people affected will increase to 578 and 700 million, respectively2. Diabetes impacts the world economy, with healthcare spending putting further strain on healthcare systems. Converting into values, approximately US$ 760 billion are invested annually in diabetic care, reaching up to US$ 845 billion in the following years3.
The diabetes development involves several risk factors, such as genetic predisposition, obesity, pathophysiological and environmental factors and the individual’s lifestyle, especially aspects of diet and sedentary lifestyle4. Numerous complications can worsen the condition of patients with diabetes, including cardiovascular and renal diseases, neuropathies, retinopathy and limb amputation, and these aggravations are mainly responsible for the increase in morbidity, disability and mortality among diabetic patients5. Thus, measures need to be taken to improve the patient’s quality of life, relieving suffering and reducing the frequency of deaths.
In this sense, due to the current scenario of diabetes and projections for the next 25 years, the concern of competent health bodies and scientists who are willing to research forms of treatment, prevention and control of complications associated with this pathology is notorious.
Polysaccharides obtained from fungi have shown biological properties that make them capable of reducing hyperglycemia. In addition, the literature reports low or absence of toxicity, low cost, rare negative side effects and a broad spectrum of associated therapeutic actions6. Microbial organisms including fungi, yeasts and algae have the advantage of availability, easy cultivation and their metabolites can be synthesized on an industrial scale in large scale bioreactors in fermentation processes7. The diversity of microorganisms is a factor that corroborates the success of fungi in biotechnology. Filamentous fungi stand out among other microorganisms, as they are easy to obtain, cultivate and manipulate8. In addition, they are relevant as a natural source to acquire biologically active substances9.
This study presents a non-systematic review that analyzes the context and potential of fungal polysaccharides for the prevention and control of diabetes. The literature lacks review works that bring together the developments of polysaccharides for the treatment of diabetes. This study sought to remedy this knowledge gap.
2 GENERAL CONSIDERATIONS ABOUT DIABETES MELLITUS
Diabetes Mellitus is one of the main syndromes of chronic evolution that affects the population worldwide and at all ages and is characterized by multifactorial disorders such as metabolic and hormonal problems. According to the Guidelines of the Brazilian Society of Diabetes10 and the American Diabetes Association11 (Standards of Medical Care in Diabetes), diabetes can be classified based on its etiology, and the most prevalent types are diabetes mellitus type 1 and type 2.
The etiology of diabetes is mainly related to a deficiency in insulin secretion or action. Insulin is a hormone secreted by pancreatic β cells that stimulates glucose uptake by tissues by causing the translocation of the glucose receptor GLUT4 from intracellular vesicles to the plasma membrane, allowing glucose to enter the cell and consequently decreasing blood glucose. Loss or deficit in pancreatic β-cell function leads to irregular insulin secretion, which causes hyperglycemia12. Figure 1 displays the basic mechanism of insulin action and the steps affected in type 1 and type 2 diabetes.
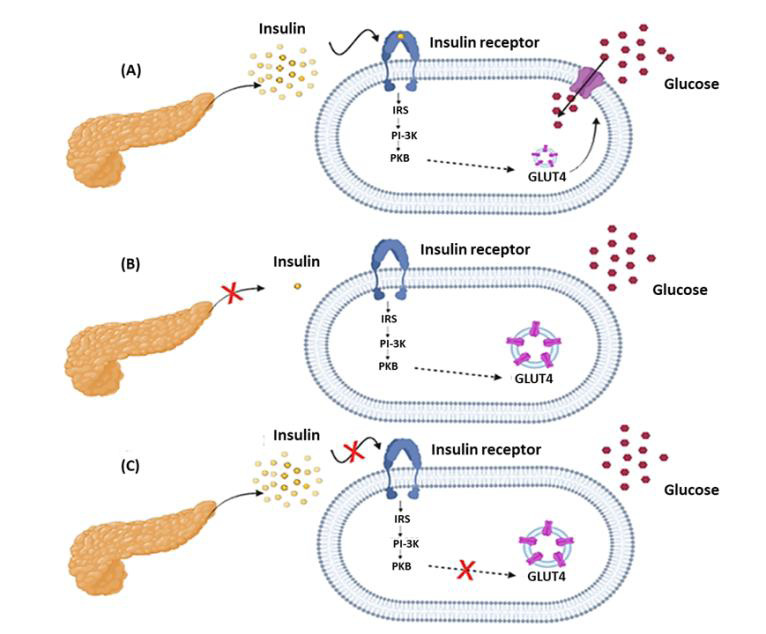
Figure 1 – Basic insulin signaling mechanism (A) and steps affected in type 1 diabetes (B) and type 2 diabetes (C).
Type 1 diabetes can be subdivided into type 1A diabetes and type 1B diabetes. In type 1A diabetes, insulin deficiency occurs due to autoimmune destruction of pancreatic β-cells, with the prevalence of autoantibodies (anti-islet, anti-insulin, anti-IA-2 and anti-GAD65) as markers of autoimmune disease. In type 1B diabetes or idiopathic, insulin deficiency occurs for an unseen reason, with the absence of markers of autoimmune disease13. Type 1A diabetes goes beyond an autoimmune disease, it is a direct relationship between environmental and genetic factors, metabolism, microbiota and the immune system of each carrier14.
Type 2 diabetes is characterized by insulin resistance, however, there may also be a gradual loss of its secretion. Type 2 diabetes has a complex and multifactorial pathophysiology, including environmental and genetic factors. Insulin resistance is associated with the ability of cytokines such as TNF-α, IL-6 and resistin to induce a state of chronic inflammation, and this metabolic dysfunction leads to insulin resistance with consequences mainly in adipose, liver and muscle tissues15.
Diabetes can also affect pregnant women, called gestational diabetes, which causes hyperglycemia to different degrees. This increase in blood glucose levels during the gestational period is mainly due to the influence of the large amounts of hormones that the pregnant woman presentes10.
Understanding the pathophysiology of diabetes is essential to take effective measures to manage and treat the disease, with a view to improving the quality of life of patients.
2.1 MANAGEMENT AND CURRENT TREATMENTS
The treatment of diabetes seeks to control blood glucose and varies according to the classification of the disease, the level at which the disease is installed in the body, and the patient’s personal factors. In addition to drug therapy, the treatment or prevention of diabetes includes lifestyle changes, such as a balanced and healthy diet, regular physical activity, smoking and alcohol cessation16.
The recommended therapy for diabetes 1 is insulin therapy, since the pancreas cannot produce the insulin needed by the body. In type 2 diabetes, to maintain normal blood glucose levels, it is possible to use different types of antidiabetic drugs.
Many of the drugs used in the treatment of diabetes have significant side effects in the life of the person with the disease. Metformin and acarbose, for example, which are among the most commonly used drugs in the treatment of diabetes, have numerous side effects on the body, such as diarrhea, flatulence and náusea17. Seeking to minimize such effects, hypoglycemic drugs from natural sources are beginning to have their place in scientific research.
3 HYPOGLYCEMIC POTENTIAL OF FUNGAL POLYSACCHARIDES
Polysaccharides are natural macromolecules in living beings. In fungi, polysaccharides can be found in the cell wall, exerting a structural function inside the cell as a source of energy reserve or in the form of exopolysaccharide. Fungal polysaccharides can present several biological activities, such as antioxidant, anticoagulant, anti-inflammatory, antimicrobial, antidiabetic, and antitumor activities18-19.
Numerous studies have reported the ability of polysaccharides obtained from fungi act as hypoglycemic agents for the treatment of diabetes. The studies grouped in Table 1 gather the mechanisms by which fungal polysaccharides may be acting and that result in antidiabetic potential, including the effects: hypoglycemic, hypolipidemic, antioxidant, anti-inflammatory, inhibition of digestive enzymes such as α-amylase and α-glucosity, in addition to the influence of IRS1 (insulin receptor substrate-1), PI3K (phosphoinositide 3-kinases) and JNK1 (c-Jun N-terminal kinase) signaling pathways (Figure 2). Studies on the hypoglycemic activity of polysaccharides should, above all, consider aspects such as the choice of the experimental model, whether in vitro or in vivo, and the elucidation of the composition and structure of these biomolecules20.
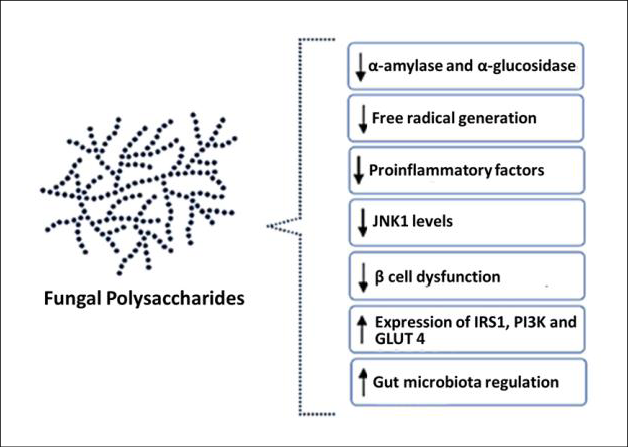
Figure 2 – Main effects of fungal polysaccharides on glucose homeostasis
Table 1 – Fungal polysaccharides and potential hypoglycemic effects
|
Fungus |
Polysaccharide Composition |
Study model |
Results |
Reference |
|
Auricularia cornea var. Li. |
ACP: mannitol (32.41%), glucuronic acid (6.96%), rhamnose (0.32%), glucose (42.35%), galactose (0.77%), xylose (16.83%) and fucose (0.36%). |
Female Kunming mice with diet-induced type 2 diabetes. |
Improvement of symptoms of hyperglycemia, reduction in blood lipid levels, reduction of liver and kidney damage and increased content of superoxide dismutase, liver glycogen and insulin. |
21 |
|
Catathelasma ventricosum |
CVP-1S: Glucose, galactose and fucose. |
Male ICR mice with streptozotocin-induced type 2 diabetes. |
Antioxidant, hypoglycemic and hypolipidemic activities. |
22 |
|
Coprinus comatus |
ICPS e IDPS: mannose, glucose, galactose, xylose and arabinose; ECPs e EDPs: mannose, galactose and xylose. |
In vitro assay for DPPH radical scavenging capacity, hydroxyl radical scavenging capacity and α-amylase inhibition. |
ICPs showed better antioxidant activity and inhibition of α-amylase activity in relation to IDPS. |
23 |
|
Cordyceps militaris |
AE-PS: fucose, ribose, arabinose, xylose, mannose, galactose and glucose. |
Mice with type 2 diabetes induced by a high-fat diet and streptozotocin. |
Decreased levels of serum lipids, lipid peroxidation and blood glucose; improvement of insulin resistance; increased activity of antioxidant enzymes; decrease in liver, kidney and pancreas lesions in mice. |
24 |
|
Cordyceps taii |
CTP: glucose, galactose and mannose (molar ratio= 1.14:1.00:1.66) |
Streptozotocin-induced diabetic male Kunming mice. In vitro assay of DPPH free radical scavenging capacity, hydroxyl and superoxide anion, reducing power and chelating capacity. |
Antidiabetic effect of CTP is associated with immunoregulation and anti-inflammatory capacity. |
25,26 |
|
Ganoderma lucidum |
F31: glucose, mannose, xylose, arabinose, galactose and ribose. |
Male Kunming mice with streptozotocin-induced type 2 diabetes. |
Decrease in levels of hepatic glucose-regulated enzyme mRNA through AMPK activation and improvement in insulin resistance. |
27 |
|
Gomphidiaceae rutilus |
AGRP: glucose (70.8%), galactose (13.2%), mannose (6.7%) and xylose (6.1%); AGRP-N: glucose (53%), galactose (29.4%), mannose (9.8%) and fucose (4.4%) and AGRP-A: glucose (69.3%), galactose (14.2%) and mannose (10.9%). |
Male C57BL/6J and ob/ob leptinam-deficient mice used as a model for obesity-induced type 2 diabetes. |
AGRP and AGRP-N reduced blood glucose and improved insulin sensitivity, in addition to inhibiting lipid deposition in the liver. |
28 |
|
Grifola frondosa |
GFP-W: L-fucose, L-xylose, D-mannose, D-glucose, D-galactose. |
HepG2 cells and male ICR mice with streptozotocin-induced type 2 diabetes. |
Antidiabetic effect by regulating the IRS1/PI3K/JNK and GLUT4 pathways. |
29 |
|
Grifola frondosa |
- |
Male Wistar rats diet-induced type 2 diabetes and Streptozotocin. |
Improved renal pathological changes, modulate serum levels of oxidant factors, inhibited NF-κB activity. |
30 |
|
Grifola frondosa |
GFP-N: L-arabinose, D-mannose and D-glucose. |
Male ICR mice with type 2 diabetes induced by streptozotocin and a diet high in sucrose and fat. |
Reduction in fasting blood glucose levels, improvement of oral glucose tolerance, alleviation of insulin resistance and protection against liver and kidney damage with reduced inflammation. Acting in the regulation of the IRS1/PI3K and JNK pathways and in the intestinal microbiota. |
31 |
|
Grifola frondosa |
GFP: mannose, rhamnose, glucuronic acid, galacturonic acid, glucose, galactose and fucose, with molar ratios of 25.49:5.18:9.49:7.30:27.59:15.02:9.92. |
Male Kunming mice with diabetes induced by a high-fat and streptozotocin diet. |
Modification of the intestinal microbiota and regulation of genes related to the hepatic metabolism of glycolipids. |
32 |
|
Inonotus obliquus |
- |
Male Kunming mice with streptozotocin-induced type 2 diabetes. |
Antihyperlipidemic and antihyperglycemic effects by activating the PI3K-Akt signaling pathway; improves insulin resistance and protects tissues from oxidative damage. |
33 |
|
Inonotus obliquus |
IOEP1: galatose e mannose; IOEP2: arabinose. |
In vitro assay with HepG2 cells and insulin resistant HepG2 cells. |
Increased glucose consumption by HepG2 cells and insulin-resistant HepG2 cells. |
34 |
|
Inonotus obliquus |
HIOP1-S: glucose and galactose; HIOP2-S: mostly glucose. |
Male ICR mice with streptozotocin-induced type 2 diabetes. |
α-glucosidase inhibitory activities and increased glucose consumption by HepG2 cells. |
35 |
|
Lachnum YM240 |
LEP: Mannose and galactose (16.3:1.0). |
Male mice with type 2 diabetes induced by streptozotocin and high-fat diet. |
Hypoglycemic and hypolipidemic effects. |
36 |
|
Pleurotus eryngii |
PEP: Glucose, mannose e galactose. |
Kunming mice with type 2 diabetes induced by a high-fat diet. |
Hypolipidemic and hypoglycemic activities. |
37 |
|
Pleurotus eryngii |
PEBP-II e PEMP-II: D-Mannose, D-Glucose, D-Galactose. |
In vitro α-glucosidase inhibition assay. |
Inhibition of α-glucosidase action by competitive inhibition. |
38 |
|
Phellinus linteus |
Glucose, arabinose, fucose, galactose and xylose. |
Sprague-Dawley rats with type 2 diabetes induced with a high-fat diet and streptozotocin. |
Increased levels of short-chain fatty acids, maintenance of intestinal barrier function and reduction of lipopolysaccharide content in the blood, helping to reduce systemic inflammation and reverse insulin resistance. |
39 |
|
Phellinus pini |
D-glucose, D-mannose, D-galactose and L-rhamnose. |
In vitro α-Glucosidase inhibition assay and insulin resistant HepG2 cell models. |
Inhibition of α-glucosidase and improvement of glucose consumption in insulin resistant HepG2 cells. |
40 |
|
Suillellus luridus |
SLPC-1S: galactose, glucose, arabinose and mannose at a molar ratio of 44.9:27.6:14.7:12.8. |
Male ICR mice with streptozotocin-induced diabetes. |
Improvement in blood glucose levels in diabetic rats, reduction in body weight, hyperlipidemia, oxidative stress and inflammation. |
41 |
To experimentally evaluate the hypoglycemic activity of a compound in an animal model, it is necessary to initially induce a diabetes condition. Most studies have used streptozotocin (STZ) as an inducer because it is a toxic agent for pancreatic β cells22,30,33,42. In its structure, STZ has a glucose molecule and a methylnitrosourea. As a mechanism of action, when STZ recognizes the GLUT2 transporter in the plasma membrane, it accumulates in the pancreatic beta cells and the methylnitrosourea portion, which has an alkylating property, is responsible for the cytotoxic effects, interfering with the production and secretion of insulin, causing diabetes43.
Fasting blood glucose, as well as the oral glucose tolerance test, are the most recommended tests for the diagnosis of diabetes11. In one study, STZ was used to increase blood glucose levels and alter the oral glucose tolerance capacity of mice. They then used polysaccharides obtained from Cordyceps militaris (AE-PS)24 as a treatment to reverse these parameters in mice with type 2 diabetes. Furthermore, polysaccharides from Grifola frondosa (GFP-W) improved oral glucose tolerance in STZ-induced diabetic mice29.
One of the criteria usually used to assess diabetes is insulin resistance. The dynamic stability of insulin levels depends on the relationship between the state of β-cells, peripheral tissues, liver and their basic state of the insulin feedback loop42. Polysaccharides obtained from the mushroom Phellinus linteus (PLPE) are able to reverse insulin resistance and suppress hepatic glucose production in rats with induced diabetes 2. PLPE was able to improve insulin resistance, reduce the state of inflammation and restore hepatic insulin signaling by inhibiting the activation of the JNK and NF-κB pathways. These pathways, when activated, lead to the inhibition of insulin signaling in the liver, with consequent insulin resistance. PLPE was able to reverse the disruption of the intestinal barrier caused by diabetes, and its potential to regulate the composition of the intestinal microbiota makes it a potential component for application as a prebiotic39.
Studies with the polysaccharide extracted from the mushroom Catathelasma ventricosum (CVP-1S) showed hypoglycemic activity analogous to that of glibenclamide. CVP-1S decreased the serum glucose levels of diabetic mice, with higher performance compared to glibenclamide. CVP-1S improved insulin levels by preventing cell damage and reducing resistance to regulate blood glucose, body weight and liver glycogen in diabetic mice22.
Diabetes is also related to disorders of lipid metabolism. The liver is the main organ that metabolizes lipids and maintains cholesterol homeostasis. Type 2 diabetes is closely related to dyslipidemia, due to the decrease in the sensitivity of the receptors on the membrane of adipose cells to insulin in this condition, causing the accumulation of fatty acids in the blood and favoring the development of diabetes42,44.
Polysaccharides extracted from Grifola frondosa and Suillellus luridus were able to improve the signs and symptoms of Sof mice with type 2 diabetes induced by STZ, regulating lipid metabolism and controlling cholesterol and triglyceride levels32,41.
Diabetes is also related to oxidative stress, the imbalance between free radicals and the body’s antioxidant capacity. Insulin resistance or deficiency in diabetic patients, in addition to decreasing glucose uptake, can increase free radical levels42. Oxidative damage to pancreatic islet cells is an important factor in diabetes22.
Antioxidant enzymes, such as superoxide dismutase, glutathione peroxidase and catalase, control the formation of reactive oxygen species and, consequently, oxidative stress. As a mechanism, superoxide dismutase can stimulate the dismutation of superoxide radicals into hydrogen peroxide and oxygen. Hydrogen peroxide, in turn, is decomposed by the action of glutathione peroxidase and catalase, preventing the formation of reactive oxygen species (ROS)22,24,44.
Studies with polysaccharides obtained from Cordyceps militaris (AE-PS) showed antioxidant potential to treat and prevent pathologies induced by ROS24. The antioxidant action of polysaccharides can be explained by their ability to donate hydrogen atoms by reducing free radicals. AE-PS improved the activity of superoxide dismutase, glutathione peroxidase and catalase in the liver, pancreas and kidneys, in addition to decreasing the concentration of malondialdehyde in STZ-induced diabetic mice. The results suggest that AE-PS can be used for the control and prevention of type 2 diabetes due to its antioxidant action in vivo24. Studies with mice treated with CVP-1S also significantly increased the activity of these antioxidant enzymes22.
Antidiabetic mechanisms can also be explained by some metabolic pathways, mainly IRS1, PI3K and JNK1 signaling. These flags along with GLUT-4 play a key role in glucose metabolism and insulin resistance. The mechanism begins with the stimulation of the intrinsic kinase activity of the insulin receptor to activate PI3K signaling, which inhibits JNK1, and finally, GLUT-4 transports glucose regulated by insulin and consequently reduces blood glucose29.
The GFP-W polysaccharide obtained from Grifola frondosa exhibited a hypoglycemic effect by increasing the expression levels of IRS1, PI3K and GLUT-4 in HepG2 cells and reducing the levels of JNK1. Regulation of such pathways by GFP-W improved insulin resistance in treated cells29. In another in vitro study, polysaccharides extracted from Inonotus obliquus (IOEP1 and IOEP2) significantly increased the rate of glucose uptake from HepG2 cells in relation to metformin33, contributing to diabetes control.
Elevated blood glucose levels can contribute to the release of proinflammatory mediators and generate a chronic inflammatory response that damages pancreatic β-cells. Destruction of β cells decreases insulin secretion, and as a consequence, hyperglycemia occurs45. Studies show that fungal polysaccharides can control the progression of type 2 diabetes by inhibiting proinflammatory factors. The CTP polysaccharide obtained from Cordyceps taii was able to significantly decrease the circulating levels of proinflammatory cytokines (IL-6, TNF-α and CRP) in diabetic mice25.
Fungal polysaccharides have the ability to act as inhibitors of important enzymes in the degradation of carbohydrates such as glycosidases and amylases46. To be absorbed and enter the circulatory system, carbohydrates ingested in the diet need to be degraded to monosaccharides. Salivary and pancreatic amylases degrade dietary polysaccharides into oligosaccharides or disaccharides. α-amylases hydrolyze the α bonds of polysaccharides. The α-Glucosidase enzymes secreted by intestinal cells hydrolyze oligosaccharides into glucose and fructose (monosaccharides) in the small intestine, enabling carbohydrate absorption47. Thus, by inhibiting these enzymes, it is possible to prevent postprandial hyperglycemia in diabetic patients.
Polysaccharides isolated from Inonotus oblíquos35 showed significant results in the inhibition of glycosidic enzymes. Furthermore, the PEBP-II and PEMP-II polysaccharides obtained from Pleurotus eryngii showed potential as α-glucosidase inhibitors in a dose-dependent manner38, while ICPS, IDPS, ECPs and EDPs polysaccharides obtained from Coprinus comatus had an inhibitory effect on α-amylase activity, reflecting a potential hypoglycemic effect23.
In addition to their role in inhibiting enzymes involved in the degradation of carbohydrates to monosaccharides, fungal polysaccharides have demonstrated their antidiabetic potential via stimulation of the intestinal microbiota48.
The intestinal microbiota performs important functions for the organism, such as protection against pathogens, metabolic functions, and immunomodulation. The alteration of the intestinal microbiota is a determining factor for the emergence of diseases, including obesity and diabetes. Fungal polysaccharides are not degraded by human digestive enzymes and are fermented in the gut, where they can improve metabolic syndrome by regulating the gut microbiota48-49.
The regulation of the intestinal microbiota was reported após o uso de the GFP-N polysaccharide from Grifola frondosa. After the administration of GFP-N in mice, it was possible to observe an increase in bacteria from the Bacteroidetes group and a lower abundance of Firmicutes and Proteobacteria. In addition, there was an increase in the prevalence of Akkermansia, Lactobacillus and Turicibacter species. The decrease in Bacteroides and the increase in Firmicutes may be related to the improvement in glucose absorption by improving insulin resistance and reducing the state of inflammation31.
The development of diabetes is generally related to a decrease in the concentration of short-chain fatty acids (SCFAs). SCFA-producing bacteria such as Blautia and Alloprevotella are important for slowing inflammation and decreasing insulin resistance50.
Studies indicate that fungal polysaccharides can contribute to the control of diabetes by acting via numerous mechanisms that culminate in the reduction of blood glucose. These biomolecules show promise for several pharmacotherapies in humans, which is mainly attributed to their safety and low toxicity18. The growing number of studies that point out the biological activities of fungal polysaccharides have been emphasizing, especially in their mechanism of action and in the structure-activity relationship19.
Some developing countries use natural polysaccharides with hypoglycemic properties as adjuvants to conventional therapies51. However, the use of fungal polysaccharides is still not widespread since clinical studies are lacking, proving their efficacy and safety in humans. Studies aimed at hypoglycemic therapy with natural products need to be stimulated, and fungal polysaccharides represent a perspective in this context, as they demonstrate important hypoglycemic properties, which make them promising as coadjuvants in the prevention and control of diabetes mellitus.
Although the results found in the studies are promising, the models used may have limitations. Despite the existence of in vitro and in silico models, animal models are still more desirable in diabetes studies, as they allow a systemic understanding of the interactions involved in the disease. Rodent models are highlighted in pathogenesis studies, while non-human primate models have translational potential due to their immunological similarity to humans. Thus, despite its limitations, the combination of experimental models is essential to optimize the translational value and understand the real potential of a given substance against diabetes52.
4 CONCLUSION
There is a growing interest in the search for drugs based on fungal polysaccharides that have resulted in hypoglycemic activities in vivo and in vitro. The studies evaluated in this work regarding the use of fungal polysaccharides for the control and prevention of diabetes mellitus are encouraging, and the main mechanisms involved are the improvement in the antioxidant capacity, resulting in less tissue oxidative damage, anti-inflammatory effects, inhibition of digestive enzymes to reduce postprandial hyperglycemia, hypolipidemic effects, regulation of IRS1, PI3K, JNK1 and intestinal microbiota signaling pathways. Such mechanisms contribute to the reduction of organ damage, such as the pancreas, liver and kidneys, and to the reestablishment of blood glucose homeostasis. Thus, fungal polysaccharides can be used as adjuncts to enhance therapy. The main limitation of the analyzed studies was the lack of clinical tests to confirm the efficacy in humans. Studies in this direction need to be stimulated.
REFERENCES
- Sun Y, Ma C, Sun H, Wang H, Peng W, Zhou Z, et al. Metabolism: A Novel Shared Link between Diabetes Mellitus and Alzheimer’s Disease. Journal of Diabetes Research. 2020;2020:1-12.
- Saeedi P, Petersohn I, Salpea P, Malanda B, Karuranga S, Unwin N, et al. Global and Regional Diabetes Prevalence Estimates for 2019 and Projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th Edition. Diabetes Research and Clinical Practice. 2019;157(157):107843.
- Williams R, Karuranga S, Malanda B, Saeedi P, Basit A, Besançon S, et al. Global and regional estimates and projections of diabetes-related health expenditure: results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Research and Clinical Practice. 2020;108072.
- Vicente-Herrero MT, Ramírez-Iñiguez de la Torre MV, Delgado-Bueno S. Diabetes mellitus and work. Assessment and questionnaires revision. Endocrinologia, Diabetes Y Nutricion. 2019;66(8):520-7.
- Jiang X, Meng W, Li L, Meng Z, Wang D. Adjuvant Therapy With Mushroom Polysaccharides for Diabetic Complications. Frontiers in Pharmacology. 2020;11.
- Korolenko TA, Bgatova NP, Vetvicka V. Glucan and Mannan – Two Peas in a Pod. International Journal of Molecular Sciences. 2019;20(13).
- Sánchez OJ, Montoya S, Vargas LM. Polysaccharide production by submerged fermentation. Polysaccharides. Springer, Cham, 2015:451-473.
- Stübiger G, Wuczkowski M, Mancera L, Lopandic K, Sterflinger K, Belgacem O. Characterization of Yeasts and Filamentous Fungi using MALDI Lipid Phenotyping. Journal of Microbiological Methods. 2016;130:27-37.
- Valasques Junior GL, Boffo EF, Santos JDG, Brandão HN, Mascarenhas AJS, Cruz FT, et al. The extraction and characterisation of a polysaccharide from Moniliophthora perniciosa CCMB 0257. Natural Product Research. 2017;31(14):1647-54.
- Brazilian Society of Diabetes (SBD). Brazilian Diabetes Society Guidelines 2019-2020. Available from: http://www.saude.ba.gov.br/wp-content/uploads/2020/02/Diretrizes-Sociedade-Brasileira-de-Diabetes-2019-2020.pdf
- American Diabetes Association. Standards of Medical Care in Diabetes-2019. Journal of Clinical and Applied Research and Education. 2019;42:1-204.
- Meyts P. The insulin receptor and its signal transduction network. Endotext, 2016.
- Cheng BW, Lo FS, Wang AM, Hung CM, Huang CY, Ting WH, et al. Autoantibodies against islet cell antigens in children with type 1 diabetes mellitus. Oncotarget, 2018;9(23):16275.
- DiMeglio LA, Evans-Molina C, Oram RA. Type 1 diabetes. The Lancet. 2018;391(10138):2449-62.
- Artasensi A, Pedretti A, Vistoli G, Fumagalli L. Type 2 Diabetes Mellitus: A Review of Multi-Target Drugs. Molecules. 2020;25(8):1987.
- Kolb H, Martin S. Environmental/lifestyle factors in the pathogenesis and prevention of type 2 diabetes. BMC Medicine. 2017;15(1).
- Razavi-Nematollahi L, Ismail-Beigi F. Adverse Effects of Glycemia-Lowering Medications in Type 2 Diabetes. Current Diabetes Reports. 2019;19(11).
- Yu Y, Shen M, Song Q, Xie J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydrate Polymers. 2018;183:91-101.
- Shi L. Bioactivities, isolation and purification methods of polysaccharides from natural products: A review. International Journal of Biological Macromolecules. 2016;92:37-48.
- Ganesan, Xu. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules. 2019;24(14):2556.
- Fu Y, Wang L, Jiang G, Ren L, Wang L, Liu X. Anti-Diabetic Activity of Polysaccharides from Auricularia cornea var. Li. Foods. 2022;11(10):1464.
- Liu Y, Chen D, You Y, Zeng S, Hu Y, Duan X, et al. Structural characterization and antidiabetic activity of a glucopyranose-rich heteropolysaccharide from Catathelasma ventricosum. Carbohydrate Polymers. 2016;149:399-407.
- Cao H, Ma S, Guo H, Cui X, Wang S, Zhong X, et al. Comparative study on the monosaccharide compositions, antioxidant and hypoglycemic activities in vitro of intracellular and extracellular polysaccharides of liquid fermented Coprinus comatus. International Journal of Biological Macromolecules. 2019;139:543-9.
- Zhao H, Lai Q, Zhang J, Huang C, Jia L. Antioxidant and Hypoglycemic Effects of Acidic-Extractable Polysaccharides from Cordyceps militaris on Type 2 Diabetes Mice. Oxidative Medicine and Cellular Longevity. 2018 Nov 25;2018:1-15.
- Liu R-M, Dai R, Luo Y, Xiao J-H. Glucose-lowering and hypolipidemic activities of polysaccharides from Cordyceps taii in streptozotocin-induced diabetic mice. BMC Complementary and Alternative Medicine. 2019;19(1).
- Xiao D, Yu S, Xiao J-H. Antioxidant activities of alkali-soluble polysaccharides from medicinal mushroom Cordyceps taii and its chemical characteristics. Biomedical Research-tokyo. 2016.
- Xiao C, Wu Q, Zhang J, Xie Y, Cai W, Tan J. Antidiabetic activity of Ganoderma lucidum polysaccharides F31 down-regulated hepatic glucose regulatory enzymes in diabetic mice. Journal of Ethnopharmacology. 2017;196:47-57.
- Yang S, Qu Y, Zhang H, Xue Z, Liu T, Yang L, et al. Hypoglycemic effects of polysaccharides from Gomphidiaceae rutilus fruiting bodies and their mechanisms. Food & Function. 2020;11(1):424-34.
- Chen Y, Liu Y, Sarker MdMR, Yan X, Yang C, Zhao L, et al. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1/PI3K-JNK signaling pathways. Carbohydrate Polymers. 2018;198:452-61.
- Kou L, Du M, Liu P, Zhang B, Zhang Y, Yang P, et al. Anti-Diabetic and Anti-Nephritic Activities of Grifola frondosa Mycelium Polysaccharides in Diet-Streptozotocin-Induced Diabetic Rats Via Modulation on Oxidative Stress. Applied Biochemistry and Biotechnology. 2019;187(1):310-22.
- Chen Y, Liu D, Wang D, Lai S, Zhong R, Liu Y, et al. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food and Chemical Toxicology: An International Journal Published for the British Industrial Biological Research Association. 2019;126:295-302.
- Guo W-L, Deng J-C, Pan Y-Y, Xu J-X, Hong J-L, Shi F-F, et al. Hypoglycemic and hypolipidemic activities of Grifola frondosa polysaccharides and their relationships with the modulation of intestinal microflora in diabetic mice induced by high-fat diet and streptozotocin. International Journal of Biological Macromolecules. 2020;153:1231-40.
- Wang J, Wang C, Li S, Li W, Yuan G, Pan Y, et al. Anti-diabetic effects of Inonotus obliquus polysaccharides in streptozotocin-induced type 2 diabetic mice and potential mechanism via PI3K-Akt signal pathway. Biomedicine & Pharmacotherapy. 2017;95:1669-77.
- Xue J, Tong S, Wang Z, Liu P. Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture. Molecules. 2018;23(12):3261.
- Liu P, Xue J, Tong S, Dong W, Wu P. Structure Characterization and Hypoglycaemic Activities of Two Polysaccharides from Inonotus obliquus. Molecules. 2018;23(8):1948.
- Wang Y, Su N, Hou G, Li J, Ye M. Hypoglycemic and hypolipidemic effects of a polysaccharide from Lachnum YM240 and its derivatives in mice, induced by a high fat diet and low dose STZ. MedChemComm. 2017;8(5):964-74.
- Chen L, Zhang Y, Sha O, Xu W, Wang S. Hypolipidaemic and hypoglycaemic activities of polysaccharide from Pleurotus eryngii in Kunming mice. International Journal of Biological Macromolecules. 2016;93(Pt A):1206-9.
- Zheng X, Sun H, Wu L, Kong X, Song Q, Zhu Z. Structural characterization and inhibition on α-glucosidase of the polysaccharides from fruiting bodies and mycelia of Pleurotus eryngii. International Journal of Biological Macromolecules. 2019;156:1512-9.
- Liu Y, Wang C, Li J, Li T, Zhang Y, Liang Y, et al. Phellinus linteus polysaccharide extract improves insulin resistance by regulating gut microbiota composition. The FASEB Journal. 2020;34(1):1065-78.
- Yang K, Jin Y, Cai M, He P, Tian B, Guan R, et al. Separation, characterization and hypoglycemic activity in vitro evaluation of a low molecular weight heteropolysaccharide from the fruiting body of Phellinus pini. Food & Function. 2021;12(8):3493-503.
- Liu Y, Liu Y, Zhang M, Li C, Zhang Z, Liu A, et al. Structural characterization of a polysaccharide from Suillellus luridus and its antidiabetic activity via Nrf2/HO-1 and NF-κB pathways. International journal of biological macromolecules. 2020;162:935-45.
- Zhang Y, Hu T, Zhou H, Zhang Y, Jin G, Yang Y. Antidiabetic effect of polysaccharides from Pleurotus ostreatus in streptozotocin-induced diabetic rats. International Journal of Biological Macromolecules. 2016;83:126-32.
- Lenzen S. The mechanisms of alloxan-and streptozotocin-induced diabetes. Diabetologia. 2008;51(2):216-226.
- Zhao H, Li S, Zhang J, Che G, Zhou M, Liu M, et al. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohydrate Polymers. 2016;151:1227-34.
- Berbudi A, Rahmadika N, Cahyadi AI, Ruslami R. Type 2 Diabetes and Its Impact on the Immune System. Current Diabetes Reviews. 2019;16(5).
- Alam F, Shafique Z, Amjad ST, Bin Asad MHH. Enzymes inhibitors from natural sources with antidiabetic activity: A review. Phytotherapy Research. 2018;33(1):41-54.
- Teng H, Chen L. α-Glucosidase and α-amylase inhibitors from seed oil: A review of liposoluble substance to treat diabetes. Critical Reviews in Food Science and Nutrition. 2016;57(16):3438-48.
- Zhao C, Yang C, Chen M, Lv X, Liu B, Yi L, et al. Regulatory Efficacy of Brown Seaweed Lessonia nigrescens Extract on the Gene Expression Profile and Intestinal Microflora in Type 2 Diabetic Mice. Molecular Nutrition & Food Research. 2018;62(4).
- Fu X, Cao C, Ren B, Zhang B, Huang Q, Li C. Structural characterization and in vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota. Carbohydrate Polymers. 2018;183:230-9.
- Zhang B, Sun W, Yu N, Sun J, Yu X, Li X, et al. Anti-diabetic effect of baicalein is associated with the modulation of gut microbiota in streptozotocin and high-fat-diet induced diabetic rats. Journal of Functional Foods. 2018 Jul;46:256-67.
- Wu J, Shi S, Wang H, Wang S. Mechanisms underlying the effect of polysaccharides in the treatment of type 2 diabetes: A review. Carbohydrate Polymers. 2016;144:474-94.
- Graham ML, Schuurman hH. Validity of animal models of type 1 diabetes, and strategies to enhance their utility in translational research. European journal of pharmacology, 2015;759:221-230.
* Graduado em Farmácia, Universidade Estadual do Sudoeste da Bahia.
† Graduando do curso de Farmácia, Universidade Estadual do Sudoeste da Bahia.
‡ Mestre em Ciências Farmacêuticas, Universidade Estadual de Feira de Santana.
§ Doutor em Biotecnologia, Universidade Estadual do Sudoeste da Bahia.
¶ Mestre em Bioquímica e Biologia Molecular, Universidade Estadual do Sudoeste da Bahia.
** Mestre em Bioquímica e Biologia Molecular, Universidade Estadual do Sudoeste da Bahia.